 'Adam and the Genome' og Doug Axes forskning om utviklingen av nye proteinfolder
'Adam and the Genome' og Doug Axes forskning om utviklingen av nye proteinfolder
Evolution news; 7. mars 2018
Bilde 1: Doug Axe, i "The Problem with Theistic Evolution," via Crossway Books.
Oversatt herfra.
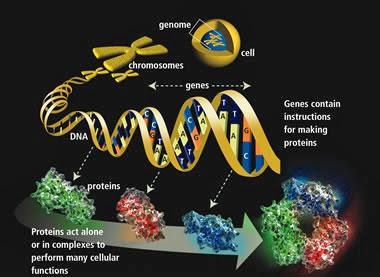
Vi har gjennomgått, i episk lengde , den litt mindre enn episke boken Adam og Genomet , av biologen Dennis Venema. Titelen på denne boken ser ut til å love en utvidet behandling av Adam, som i Adam og Eva, og det menneskelige genomet. Men gjentatte ganger trekker Venema inn andre saker, irrelevant for hvorvidt Adam og Eva eksisterte.
På enda en utflukt kritiserer han undersøkelsen til Douglas Axe, en proteinforsker som har publisert arbeid om sjeldenhet av nye proteinfolder ved å undersøke beta-laktamase-enzymer. Publisert i Journal of Molecular Biology , Axe fant  at bare ca 1 av (10 opphøyd i 77) sekvenser produserer den stabilt foldede strukturen som trengs for beta-laktamase til å fungere.
at bare ca 1 av (10 opphøyd i 77) sekvenser produserer den stabilt foldede strukturen som trengs for beta-laktamase til å fungere.
Bilde 2. Bokas forfatter: mål for Venemas angrep
Men Venema hovedmål i denne delen er egentlig ikke Doug Axe. Det er Stephen Meyer. Spesielt går Venema etter Meyers bruk av Axes forskning, og hevder at Axes arbeid bare gjelder utviklingen av beta-laktamase og ikke evolvbarheten av proteiner generelt. Han hevder at Meyer over-ekstrapolerer implikasjonene til Axes arbeid:
"Nå er disse resultatene ikke kontroversielle for Axes utviklede beta-laktamase. Det som er kontroversielt, er imidlertid Meyers påstand om at disse resultatene gjelder for utviklingen av proteiner generelt. ... Den gjennomsnittlige legmannen som leser Meyers verk, kan imidlertid bare ta ham på hans ord at forskere har konkludert med at funksjonelle, foldede proteiner generelt er svært sjeldne og dermed enige i hans vurdering at de ikke kan produseres av naturlige mekanismer." ( Adam and the Genome , s. 83-84)
Venema skriver at "Hvis forskere kunne observere en slik begivenhet [utviklingen av en nytt proteinfold], så ville det tyde på at Axes matte (og Meyers bruk av det) ikke er et pålitelig estimat for utbredelsen av funksjonelle proteinfolder." Av å forsøke å splitte "Axes matte" fra "Meyers bruk av det", synes Venema å bety at Axes argumenter er relativt sunt (siden Axe formodentlig begrenset sine konklusjoner til den konkrete utviklingen av beta-laktamase), mens Meyers ekstrapolering til det generelle tilfellet er det virkelige problemet, som bedriver et misbruk av Axes artikkel.
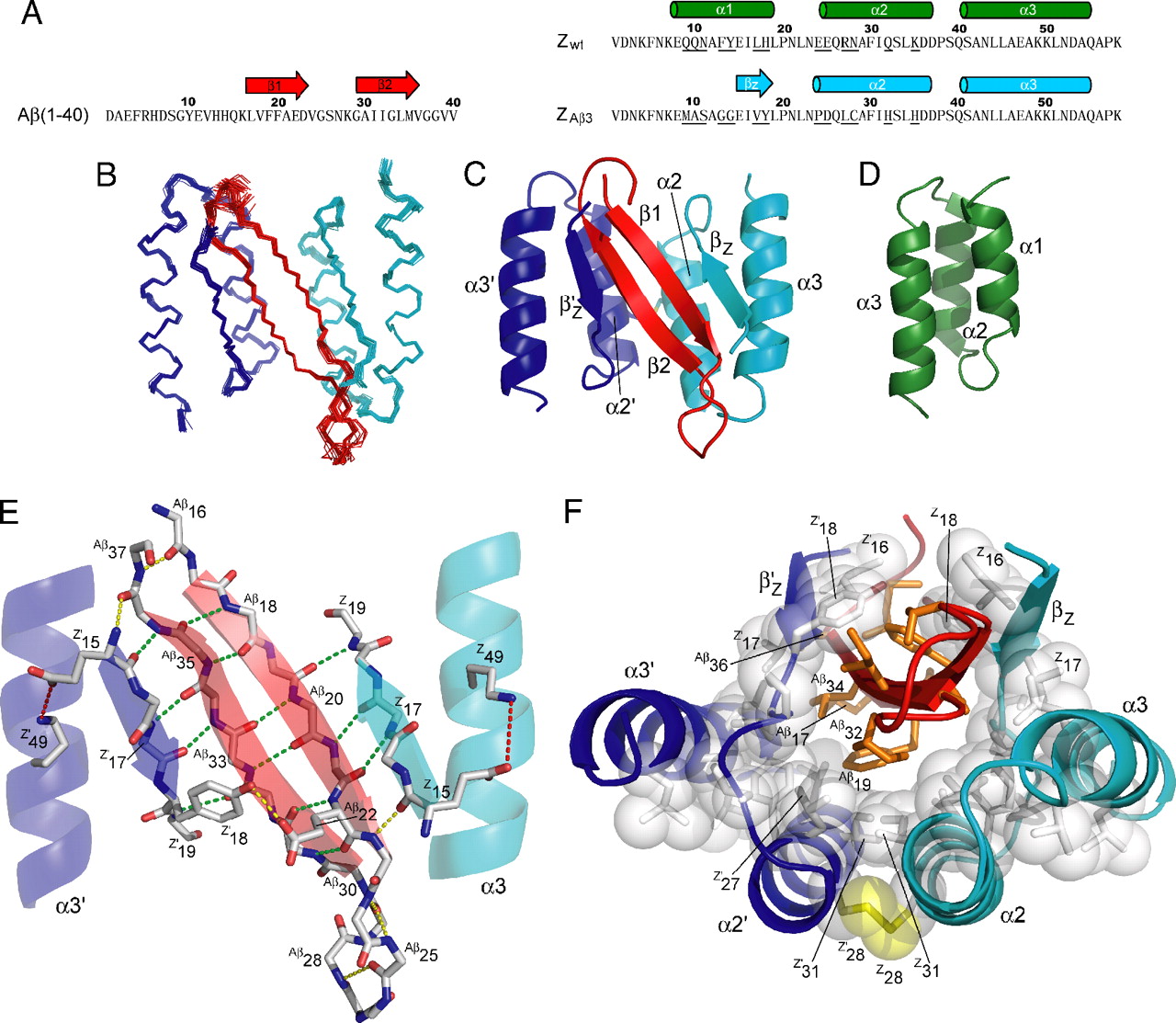
 Bilde 3. Eks. på 3D-proteinstruktur
Bilde 3. Eks. på 3D-proteinstruktur
Her gjør Venema en kritisk feil: Tilsynelatende forstår han ikke at Axes fagfelle-vurderte artikkel uttrykkelig argumenterer for å begrunne å utvide sine resultater på beta-laktamase mutagenese eksperimenter til sjeldenhet av nye proteinfolder generelt - ikke bare til opprinnelsen av beta -lactamase. Dette er ikke noe overraskende. Mange artikler om proteinstruktur, funksjon og evolvabarhet vurderer spesifikke tilfeller og drøfter de generelle implikasjonene. Meyer misbrukte ikke Axes artikkel. Meyer gjentok rett og slett de direkte konklusjonene av en artikkel som ble publisert i den prestisjetunge hovedstrøms biologiske journalen.
Tittelen til Axes artikkel, "Estimating the Prevalence of Protein Sequences Adopting Functional Enzyme Folds,"-her, tyder på at den er rettet mot å diskutere et generelt resultat. I de første ordene i Abstraktet indikerer Axe at hans resultater kan gjelde for "overordnet utbredelse av sekvensering ved bruk av funksjonelle folder":
"Proteiner benytter et bredt utvalg av bretter for å utføre sine biologiske funksjoner. Hvordan blir disse foldene først oppnådd? Et viktig skritt mot å svare på dette er å skaffe et estimat av den generelle forekomsten av sekvenser som tar i bruk funksjonelle folder."
(Douglas D. Axe, "Estimering av prevalensen av proteinsekvenser ved å vedta funksjonelle enzymfold", Journal of Molecular Biology , 341: 1295-1315 (2004).)
Bilde 4. Protein med alpha- og beta-heliks
 Ved å lese videre i artikkelen ser vi at Axe bevisst valgte å jobbe med beta-laktamase fordi den har en fold av typisk kompleksitet, og kan dermed tjene som "et modellsystem for å vurdere kravene til funksjonell dannelse av en moderat kompleks fold". Axe forklarer :
Ved å lese videre i artikkelen ser vi at Axe bevisst valgte å jobbe med beta-laktamase fordi den har en fold av typisk kompleksitet, og kan dermed tjene som "et modellsystem for å vurdere kravene til funksjonell dannelse av en moderat kompleks fold". Axe forklarer :
"Den større av de to domenene som danner beta-laktamaser av klasse A-variasjon (videre: 'det store domenet') brukt som et modellsystem for å vurdere kravene til funksjonell dannelse av en fold av moderat kompleksitet. Selv om det hovedsakelig består av alfa- helikser, inneholder dette domenet små ark regioner og signifikant sløyfe-struktur som sammen med sin størrelse (litt over 150 aminosyrer) gjør dens kompleksitet mer representativ for kjente domene-foldinger. Et annet typisk trekk ved domener, evnen til å danne spesifikke foreninger med andre domener, er sikret ved plasseringen av beta-laktamase-aktivitetsspalten i grensesnittet mellom de store og små domener."
I slutten av artikkelen ekstrapolerer Axe sine resultater fra beta-laktamase enzymer til det generelle tilfellet:
"Det er ikke åpenbart at fold-mangfoldet er like lett forklart som sekvensdiversitet, hvis funksjonelt foldede sekvenser er like sjeldne som denne analysen indikerer. En allment akseptert visning er at nye folder er sammenstilt fra små deler av eksisterende foldinger. Men i den utstrekning at en ny fold virkelig er ny, må formasjonen kreve felles løsning av minst et betydelig antall nye lokale stabiliserings-problemer av den type som er beskrevet ovenfor. Hvor sannsynlig er det at sekvenser som bærer hydropatiens underskrifter av andre folder og gir felles løsninger på stabiliseringsproblemene for disse foldene, kan bli satt sammen på en slik måte at de tilfredsstiller et nytt sett med begrensninger, like krevende, men vesentlig forskjellige? Analysen gitt her, med tanke på usikkerhetene, krever nøye undersøkelse av slike sammensetnings scenarier."
Dette er bare en teknisk måte å si at resultatene tyder på at nye folder er svært vanskelig å produsere ved naturlig utvalg. Hans eksperiment satte ut til å måle følsomheten til de lokale stabiliseringsproblemene for forstyrrelse, og dermed føles følsomheten av beta-laktamasen til destabilisering som helhet. Når proteiner er destabilisert, mister de funksjon. Dette tap av funksjonen gir et mål på sjeldenheten av stabile funksjonelle folder. Fra artikkelens sammendrag:
"Kombinert med den estimerte utbredelsen av plausible hydropatiske mønstre (for en hvilken som helst fold) og av relevante folder for bestemte funksjoner, betyr dette at den generelle forekomsten av sekvenser som utfører en spesifikk funksjon ved hvilken som helst domenestørrelse, kan være så lav som 1 av (10 opphøyd i 77) , idet det legges til bevis for at funksjonelle folder krever svært ekstraordinære sekvenser."
Hva alt dette betyr er at Meyer aldri misbrukte Axes artikkel. Han omstilte bare Axes egne peer-reviewed konklusjoner.
Bilde 5. U-sving
 Å vende retningen er fair play
Å vende retningen er fair play
Snu nå logikken rundt. Venema bruker i sin bok gjentatte ganger spesifikke eksempler på proteiner som han mener er evolverbare for å insistere på at nye komplekse protein-konstruksjoner generelt er evolverbare. Han argumenterer ofte fra det spesifikke til det generelle, og innrømmer stilltiende at Axe (og Meyers) form for argument er legitimt. (Merk, som vi så tidligere , har Venema imidlertid ikke vist at de spesifikke proteinene han diskuterer er evolverbare, og han har heller ikke vist opprinnelsen til noen komplekse egenskaper. Hans spesifikke eksempler støtter ikke hans generelle argument om at komplekse egenskaper kan utvikle seg.)
Poenget er at det er helt legitimt i vitenskap å bruke ett tilfelle som en modell for analyse av et generelt problem. Dette blir gjort hele tiden. Axes generalisering av resultater følger tradisjonen med mange lignende artikler, som kom til lignende konklusjoner om sjeldenhet av funksjonelle proteinsekvenser, og anvendte deres resultater i stor grad. For eksempel:
*Reidhaar-Olson og Sauer 1990 -her (publisert i tidsskriftet Proteiner ), muterte A-repressoren i coli og fant at bare en til (10 opphøyd i 63) sekvenser gir en funksjonell repressorfold. De generaliserte implikasjonene av deres resultater for hvordan vi forutsier proteinstruktur i andre tilfeller, ved å skrive: "Det høye nivået av degenerasjon involvert i proteinfolding antyder at de mest fruktbare tilnærmingene til strukturprediksjon vil konsentrere seg om de rester som er informasjonsrike."
*Yockey 1977 -her (publisert i Journal of Theoretical Biology ) beregnet at sannsynligheten for å generere en funksjonell cytokrom c-sekvens er én til (10 opphøyd i 65). Han generaliserte dette resultatet ved å konkludere med at mange proteiner ikke er evolvable, og til og med konkluderte med at standardmekanismene for abiogenese ikke kunne produsere slike egenskaper på en rimelig tidsplan. Han skrev at "tro på nåværende aksepterte scenarier av spontan biogenese er basert på tro, i motsetning til konvensjonell visdom."
 Bilde 6. Axe forsket også på Kbl og BioF
Bilde 6. Axe forsket også på Kbl og BioF
*Hayashi et al. 2006 -her (publisert i PLOS ONE ) fastslo at (10 opphøyd i 70) forsøk ville være nødvendige for å erverve vill-type-funksjonen av lite g3p-kappeprotein fra fd- bakteriofagen. De generaliserte deres utledede egnethets-resultater til andre tilfeller og skrev: "Landskapsstrukturen har en rekke implikasjoner for den første funksjonelle evolusjonen av proteiner og for molekylær evolusjonær ingeniørfag." Men fordi å nå høyere fitness-nivåer krevde å klare av mye brattere fitness-funksjoner ( det vil si at funksjonelle sekvenser var svært sjeldne), og dermed konkludere de som en generell sak: "I molekylær evolusjonær ingeniørvirksomhet er større biblioteks-størrelse generelt gunstig for å nå høyere stasjonær fitness."
Så Axes artikkel generaliserer sine resultater lik tidligere artikler har gjort. Faktisk rapporterer han at en artikkel fant sannsynligheten for at en sekvens som genererte korismat-mutase, var mye høyere - (10 opphøyd i -40) - enn resultatet av (10 opphøyd i -64) for beta-laktamase. Imidlertid bemerker han at "det er ingen grunn til å tro at de to estimatene er inkonsistente" siden folden som studeres av korismat-mutase artikkelen, var en mye enklere form for fold. Axe skriver at "det er viktig at enzym-folder av mer typisk kompleksitet undersøkes", slik han utformet sin studie.
Dermed rettferdiggjør Axe ekstrapoleringen av resultatene sine til andre proteiner ved å merke tilsvarende nivå av kompleksitet til det vi vanligvis ser:
"Selv om det hovedsakelig består av a-helikser, inneholder dette domenet små ark områder og en signifikant sløyfestruktur som sammen med dens størrelse (litt over 150 aminosyrer) gjør dens kompleksitet mer representativ for kjente domene-folder.
Forover og bakover
Axe er klar over Venema's type av kritikk og adresserer dem i sin artikkel. Fra begynnelsen observerer Axe at det finnes to typer mutagen-forsøk:
1. "Framover" -studier, som starter med et bibliotek av tilfeldige sekvenser, og deretter forsøker å avgjøre om det finnes noen som har funksjonene eller egenskapene til funksjonelle proteiner.
2. "Revers" -studier, som starter med et naturlig funksjonelt protein og deretter muterer det for å bestemme toleransen for endringer.
Axe påpeker at begge typer eksperimenter har fordeler og ulemper. Type (1) ("fremover") eksperimenter mislykkes vanligvis i å produsere sekvenser som tydelig ligner naturlige proteiner. Men Type (2) ("revers") eksperimenter "kan mislykkes i å ta hensyn til sekvenser som har de relevante funksjonsegenskapene i en svært rudimentær form."
Venema synes tilsynelatende at Axe utfører naive type (1) eksperimenter og dermed gjør den samme kritikken av Type (2) eksperimenter som Axe selv gjør, som om Axe ikke var oppmerksom på kritikken og ikke hadde adressert det. Venema skriver således:
"Axes eksperiment starter med et definert protein og modifiserer det. Problemet her er at det finnes andre beta-laktamaser som ikke ligner den ene Axen valgte som hans bakterio- fag. Disse andre proteiner fungerer som beta-laktamaser med en aminosyresekvens som er veldig forskjellig fra test-enzymet. Så det var sikkert sekvenser i forsøket som kunne fungere som en beta-laktamase, men ikke som en del av beta-laktamase konstruksjonen Axe valgte. Hans eksperiment ville med andre ord savne sekvenser selv for den aktuelle funksjonen." ( Ad am and the Genome , s. 84)
am and the Genome , s. 84)
Bilde 8. Passform enzym og substrat må matche
Men som vi allerede har sett, er Axe vel kjent med disse problemene, for han diskuterer dem og utformet til og med sine eksperimenter for å imøtekomme dem. Axe skriver i sin artikkel:
"Siden mange forskjellige folder kan være sammenlignbare for enhver gitt enzymatisk funksjon, er det viktig at vi har noen måte å faktorere dette inn. Med andre ord, hvis forekomsten av sekvenser som utfører en bestemt funksjon enzymatisk, er vår primære interesse, da må ikke vår analyse anta nødvendigheten av en bestemt fold."
Faktisk var Axes mutagenese-eksperimenter spesifikt tilpasset det faktum at andre beta-laktamaser kan ha svært forskjellige sekvenser. Han skriver:
Ved å benytte sekvensinformasjon fra en rekke relaterte beta-laktamaser, er det mulig å rette analysen av denne enkelt-folden på en slik måte at den belyser de viktigste aspektene av sekvensfunksjons-forholdet som må undersøkes for å kunne vurdere den samlede utbredelse av enzymatisk funksjon.
Axe tilpasset en tilnærming som ligner på en tidligere mutagenese-studie av korismat (chorismate). Etter å ha fortalt om noen av problemene som står overfor Type (2) eksperimenter, skriver Axe:
"Hvordan kan de andre vanskelighetene unngås? En nylig studie av kravene til korismat mutase-funksjon i laboratoriet demonstrerer en lovende tilnærming. Korismat-mutasegen-biblioteker fremstilt i dette arbeidet var begrenset til å bevare alle residuer på aktivt sted og sekvensiell oppstilling av hydrofobe og hydrofile sidekjeder tilstede i en naturlig versjon av enzymet. Innenfor disse begrensningene var imidlertid spesifikke rest-tilordninger i det vesentlige tilfeldige, hvilket resulterte i mange forstyrrende forandringer gjennom de kodede proteiner. Dette er et eksempel på omvendt tilnærming, ved at den bruker en naturlig sekvens som utgangspunkt, men fordi de produserte varianter har omfattende forstyrrelser i hele strukturen i stedet for bare lokal forstyrrelse, gir de pålitelig informasjon om stringentheten til funksjonelle krav."
Bilde 9. Protein.syntase med funksjonell form
 Axes mutagenese-forsøk på beta-laktamase følger en lignende tilnærming:
Axes mutagenese-forsøk på beta-laktamase følger en lignende tilnærming:
"Som i korismat mutase-studien, vil forstyrrende substitusjoner i hele det store domenet gi en marginalt tilstrekkelig sekvens-kontekst for å vurdere kravene til lavnivåfunksjon."
Dermed er det Venemas kritikk av Axes eksperimentelle design som kommer til kort, ikke Axes eksperimenter.
Vanlige og falske innvendinger
Men de vanligste (men falske) innvendingene som Axe rutinemessig står overfor, vises også i Venema's bok. Venema skriver:
"For det første, husk at Axes eksperiment brukte et mutert, knapt funksjonelt protein fordi det er velkjent at naturlige proteiner er i stand til å tolerere mange mutasjoner uten å miste funksjonen. Det er ikke kontroversielt å si at Ax hadde brukt et normalt, stabilt protein, han ville ha funnet langt mer "funksjonelle" proteiner i sitt eksperiment." (Adam and the Genome , s. 83-84)
Dette er en misforstått kritikk. Axe ønsket ikke å måle hvor mange mutasjoner et villtype protein kunne tolerere. Han ønsket å bestemme hvor vanskelig det er å gå fra en ikke-funksjonell, men signatur-kompatibel fold til en funksjonell. Med andre ord spurte han om grensevilkårene for å ha en funksjonell fold - hvilken andel proteiner med riktig hydrofob/ hydrofil profil var faktisk funksjonell? Dette kan bare måles ved grensen mellom de to forholdene.
For det andre beklager Venema at Axe muterte proteiner i grupper.
Bilde 10. Årsaker til punktmutasjoner
 "Husk også at Axe erstattet flere aminosyrer om gangen i testene. Det ville rett og slett ikke være tid til å prøve alle mulige single mutasjoner, dobbel mutasjon, triple mutasjon og så videre. Mange av de små mutasjonene ville ikke forventes å fjerne restfunksjonen til det konstruerte proteinet også. Evolusjon, som vi har sett, fungerer vanligvis via enkle mutasjoner, ikke mange samtidige. Axes eksperiment trengte å bruke flere samtidige mutasjoner for å redusere antall prøver, men dette betyr at oppsettet hans er mindre relevant for spørsmålet om hvordan evolusjonen kan bygge nye proteinstrukturer over tid. Faktisk forventer biologer at samtidige mutasjon av flere aminosyrer vil redusere funksjonen - spesielt for et protein som allerede er mutert til det punktet som knapt fungerer i utgangspunktet." (s. 84)
"Husk også at Axe erstattet flere aminosyrer om gangen i testene. Det ville rett og slett ikke være tid til å prøve alle mulige single mutasjoner, dobbel mutasjon, triple mutasjon og så videre. Mange av de små mutasjonene ville ikke forventes å fjerne restfunksjonen til det konstruerte proteinet også. Evolusjon, som vi har sett, fungerer vanligvis via enkle mutasjoner, ikke mange samtidige. Axes eksperiment trengte å bruke flere samtidige mutasjoner for å redusere antall prøver, men dette betyr at oppsettet hans er mindre relevant for spørsmålet om hvordan evolusjonen kan bygge nye proteinstrukturer over tid. Faktisk forventer biologer at samtidige mutasjon av flere aminosyrer vil redusere funksjonen - spesielt for et protein som allerede er mutert til det punktet som knapt fungerer i utgangspunktet." (s. 84)
Her synes Venema helt å ha gått glipp av poenget med Axes eksperiment. (Han er ikke den eneste.) Axe prøvde ikke å bestemme hvordan man skal bygge et protein. Han målte ikke hvordan evolusjonen fungerte. Han ville bare se hvilken andel muterte proteiner som var stabile nok til å utføre en beta-laktamase-reaksjon. For å gjøre dette målte han hvor mange proteiner som kunne fungere etter at en liten assosiert gruppe av aminosyrer ble endret i dem. Siden proteinfoldene stabiliseres av et nettverk av sidekjedens interaksjoner, bør destabilisering av en liten del av nettverket gi en nøyaktig avlesning om hvor kritisk regionen er for stabilitet. Gruppene ble randomisert, samtidig som de beholdt nøkkelhydrofobe og hydrofile residuer, og dermed ga den beste muligheten til å bevare den samlede strukturen. Når resultatene for hver av fire grupper ble bestemt, ble et estimat for hele domenet beregnet. I hovedsak bestemte eksperimentet terskelen for en stabil funksjonell fold som var i stand til å utføre beta-laktamasase-reaksjonen - lik det å krype ut av et ikke-funksjonelt hav til den funksjonelle bredden. Om noe, ville bruken av flere mutasjoner gi et løft til overgangen.
Bilde 11. Flerfoldige dimensjoner i proteiner
 Til slutt er det verdt å ta en titt på Venemas avsluttende ord i hans del om Axes forskning:
Til slutt er det verdt å ta en titt på Venemas avsluttende ord i hans del om Axes forskning:
"Hvis forskere kunne observere en slik begivenhet [den mekaniske opprinnelsen til et nytt proteinfold], ville det indikere at Axes matte (og Meyers bruk av den) ikke er et pålitelig estimat for utbredelsen av funksjonelle proteinfeller. Interessant nok, er det mange kjente tilfeller av akkurat dette, selv om Meyer ikke ser ut til å være klar over dem, eller om de konsekvensene de holder for hans argumentasjonslinje. La oss undersøke noen i detalj. "(s. 84-85)
Hoved-eksemplet Venema gir da er opprinnelsen til nylonase, som, som vi allerede har sett i denne serien -her, ikke er et eksempel på den spontane utviklingen av en ny proteinfold. Venema mistolket dataene.
Venema nevner også opprinnelsen til p24-2, selv om det ikke er tegn på at p24-2 involverte opprinnelsen til en ny proteinfold.
Det som er veldig merkelig er at Venema rammer hele diskusjonen som nok et forsøk på å svekke Meyers troverdighet. Han skriver, "Meyer ser ikke ut til å være klar over" av de angivelig mange eksemplene som viser at nye proteiner utvikler seg. Men i sin 2013-bok Darwins Doubt bruker Meyer et helt kapittel som går over påståtte eksempler på utviklingen av nye gener som ligner på nylonase og p24-2 . I kapittel 11, "Tenk deg et gen", undersøker Meyer følgende:
*Opprinnelsen til et frostvæske-protein hos fisk i Antarktis .
*Opprinnelsen til RNASE1B, et fordøyelsesprotein i colobin-aper.
*Opprinnelsen til Cid , et gen som koder for et histon-protein i bananfluer.
*Opprinnelsen til FOXP2 , et gen som er involvert i å regulere hjernens utvikling i forskjellige pattedyr.
Bilde 12. I genomet: 4 dimensjoner (minst)
 På denne måten vurderer Meyer praktisk talt alle standardmekanismer som påberopes for utvikle nye gener, inkludert gen-duplisering, ekson-skyfling, reposisjonering av mRNA, lateral genoverføring, mobile genetiske elementer, gen-fisjon/fusjon og de novo gen-opprinnelse. Han ignorerer ikke litteraturen om utvikling av nye gener, og diskuterer om like mange eksempler på genevolusjon i boken 'Darwins Doubt' som Venema diskuterer i 'Adam and the Genome'.
På denne måten vurderer Meyer praktisk talt alle standardmekanismer som påberopes for utvikle nye gener, inkludert gen-duplisering, ekson-skyfling, reposisjonering av mRNA, lateral genoverføring, mobile genetiske elementer, gen-fisjon/fusjon og de novo gen-opprinnelse. Han ignorerer ikke litteraturen om utvikling av nye gener, og diskuterer om like mange eksempler på genevolusjon i boken 'Darwins Doubt' som Venema diskuterer i 'Adam and the Genome'.
Venema er ikke bare uenig med Meyers argumenter. Han vil male Meyer som uvitende, inkompetent og upålitelig. Men den relevante vitenskapen er på Meyers side, ikke Venemas. Dette er selvsagt bortsett fra spørsmålet om hva Steve Meyer i det hele tatt gjør i en bok om eksistensen av Adam og Eva, et emne han (Meyer) ikke engang skriver om.
Oversettelse og bilder ved Asbjørn E. Lund
{kind=link}